
双脱氧法或酶法利用DNA 聚合酶合成单链DNA 模板的互补拷贝,这一方法最先由F. Sanger 及其合作者发展而来。DNA 聚合酶不能起始DNA 链的合成,而是在复性于“模板“DNA 的引物的3 ‘端上进行链的延伸(图1) 。链的延伸是在引物生长端的3′ 羟基掺入脱氧核糖核苷酸。双脱氧测序法利用了DNA 聚合酶能以2′, 3’-双脱氧核糖核苷酸(ddNTP) 为底物的特性。当ddNTP 被掺入到延伸着的引物的3′ 端时,由于链上3′ 羟基的缺乏,链的延伸就会终止。在4 个测序反应的每个反应中加入4 种可能的ddNTP 中的一种,即可产生4 种不同的反应。调整每个测序反应中的ddNTP 与dNTP的比例,使部分引物延伸链分别终止于每一个在模板DNA 出现该碱基的位置上。这种测序方式,每个延伸反应的产物是一系列长短不一的引物延伸链,它们都具有由复性引物所决定的固定的5’端和终止于某一ddNTP 的不定的3’ 端。
常用的双脱氧测序法有两种常用方案。最早期的双脱氧法,称之为Sanger 法(Sanger et al. 1977, 1980) , 是利用大肠杆菌DNA 聚合酶I 的Klenow 片段而发展起来的 。引物在需要测序的单链DNA 模板的3′ 端进行复性(图1) 。分成4份进行反应,每一份反应中都含有DNA 聚合酶、3 种未标记的dNTP、一种标记的dNTP 及其相应的ddNTP (图1 右侧) 。引物的延伸和标记进行至摄入ddNTP 后被终止。在接下来的反应中,追加高浓度的4 种dNTP 使未被ddNTP 终止的链再延伸
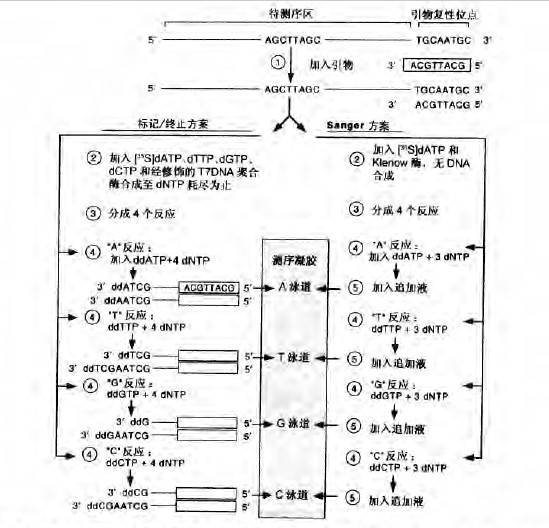
(右图), 加人Klenow 酶和放射标记的dATP (步骤2) ,然后,分成4 份进行反应(步骤3),分别加入其余的3 种dNTP 及一种ddNTP (步骤4) 。DNA 的合成进行至摄入ddNTP 后被终止。追加dNTP (步骤5) 使未被终止的链再延伸以产生更高分子质址的DNA。在“标记/终止法”中(左图) ,紧接着步骤1 ,加入限量的4 种dNTP (其中一种为放射性标记)和测序酶(步骤2 ), DNA 的合成至dNTP 耗竭为止。反应混合物等分成4 组(步骤3) ,分别加入4 种dNTP 和一种ddNTP(步骤4 ) ,恢复合成反应,但当掺入ddNTP 后,反应被特异性地终止。在每种方法中,反应终止后,样品加于测序胶的相邻泳道上,进行电泳分离。
以产生更高分子质量的DNA, 这种DNA 链滞留在测序胶的顶部无法分辨。测序产物的平均长度通过ddNTP/dNTP 的比率来控制,比率越高产物越短。
标记/终止法利用修饰的T7 噬菌体DNA 聚合酶得到进一步发展。在两个独立的反应中分别进行引物的标记和双脱氧核苷酸的掺入(图 1 左侧)。复性的引物在4 种低浓度dNTP(其中1 种是放射性标记的)存在时进行延伸。DNA 的合成持续到一种或多种dNTP 被耗竭为止,这样可保证掺入全部的标记的脱氧核糖核苷酸。标记的混合物分到4 个独立的反应中,每个反应除了含有4 种dNTP 外,还各含4 种ddNTP 中的1种。在链终止反应步骤中,高浓度的dNTP 保证DNA 逐次合成至生长链因ddNTP 的掺入而终止。测序产物的平均链长取决于标记反应中dNTP 的浓度(浓度越高产物越长)和终止反应中ddNTP : dNTP 的比例。
当使用测序酶的时,标记/终止法能得到平均长度长于Sanger 法的测序产物。所以这种方法对于从每个模板上获得最大散序列的信息是更有利的。对于测定小片段DNA(如证实结构), Sanger 法通常就足够了。如确定引物后几个核苷酸的序列信息, Sanger 法则更可靠。热稳定的DNA 聚合酶也可以用于双脱氧测序。因为耐热DNA 聚合酶可以在高温下工作,可以用它进行热循环反应,这样可以提高测序产物的产量,因而提高了灵敏度。此外,热稳定聚合酶可以使DNA 模板的二级结构变性,而DNA 的二级结构会干扰延伸进程。